光合成の仕組みは研究が進むにつれ化学的にも解明されることが多くなりました。
反応は段階的に行われるので難しく見えますが、全体を通した化学反応式で見ておくと少しは分かり易くなります。
反応系の一部、カルビン・ベンソン回路と呼吸との関係についてもみておきましょう。
チラコイドにおける反応
葉緑体のチラコイド膜の上には、光化学系Ⅰと光化学系Ⅱと呼ばれる2つの光エネルギーを捕集する反応系があります。
この2つの反応系はクロロフィルa、クロロフィルb、カロテノイドなどの光合成色素とタンパク質の複合体からできていて、カロテノイドなどの色素で集められた光エネルギーは、反応中心であるクロロフィルに集められて利用されます。
光エネルギーを受け取った光化学系Ⅱではクロロフィルから電子が飛び出して電子伝達系に流れます。
このときに電子が不足するような場合、
水( \(\mathrm{H_2O}\) )を酸素( \(\mathrm{O_2}\) )と水素イオン( \(\mathrm{H^+}\) )に分解したときの電子( \(\mathrm{e^-}\) )で補充されます。
光合成の反応全体では12分子の水が分解されます。
\( \mathrm{12H_2O\rightarrow 6O_2+24H^++24e^-}\)
ただし、後で詳しく書きますがこの分解される水分子と生成物の水とは出所が違いますので注意しておいてください。
またこのとき発生する酸素は気孔を通じて排出されます。
光エネルギーを受容した光化学系Ⅰで電子受容体にわたされた電子は、
脱水素酵素の補酵素 \(\mathrm{NADP^+}\) にわたされて \(\mathrm{H^+}\) とともに \(\mathrm{NADPH}\) となります。
このとき不足した電子は、電子伝達系を通ってきた電子で補充されるのです。
光合成で使われる電子は緑葉の場合、光化学系Ⅱの電子の補充に水の分解による電子が使われますが、
光合成細菌の場合は硫化水素( \(\mathrm{H_2S}\) )から電子が補充されます。
電子が光化学系Ⅱから光化学系Ⅰに伝達される過程を電子伝達系といいますが、
これは呼吸におけるミトコンドリアの内膜にある電子伝達系と似た仕組みです。
この電子伝達系で電子が受け渡しされるときに生じるエネルギーを利用し、
水素イオンがストロマ側からチラコイドの内側に運ばれます。
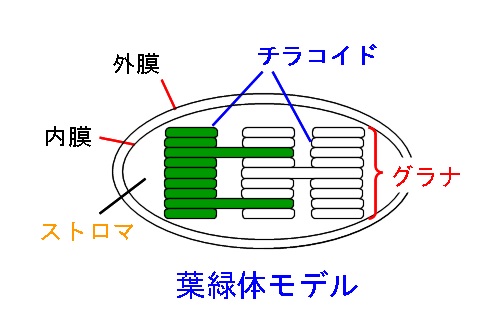
そしてチラコイド内の水素イオン濃度が上がり、ストロマ側との濃度勾配(濃度の差)が大きくなると、チラコイド膜にある膜貫通タンパク質であるATP合成酵素を通って水素イオンがストロマ側にもどります。
このときの水素イオンの流れによってATPが合成されるのですが、この反応を『光リン酸化』といいます。
光リン酸化の仕組みは、ミトコンドリアで行われる呼吸のときのATP生成に似ていますが、呼吸のときのATP合成は『酸化的リン酸化』と呼ばれます。
ストロマにおける反応(カルビン・ベンソン回路)
ストロマ部分ではチラコイド膜でつくられたATPとNADPHを使って二酸化炭素( \(\mathrm{CO_2}\) )を還元して有機物に合成する。
\(\mathrm{CO_2}\) は \(\mathrm{C_5}\) 化合物のリブロース二リン酸(RuBP)と結合し、
\(\mathrm{C_6}\) 化合物になり、すぐに分解して、
2分子の \(\mathrm{C_3}\) 化合物のホスホグリセリン酸(PGA)になります。
このときはたらく酵素は『ルビスコ』(RubisCo)といいます。
PGAはATPエネルギーとNADPHを使って還元されて、
\(\mathrm{C_3}\) 化合物のGAP(グリセルアルデヒドリン酸)となります。
この一部分は有機物(グルコース)合成に回りますが、残りはRuBPにもどります。
\(\mathrm{6\color{red}{RuBP} + 6CO_2 ⇒ 12PGA ⇒ 12GAP ⇒ 6\color{red}{RuBP}}\)
このストロマで行われる反応系を「カルビン・ベンソン回路」といいます。
「カルビン回路」と呼ばれることもありますが「カルビン・ベンソン回路」とは同じものです。
これまでの光合成の反応をまとめると
\(\mathrm{6CO_2\,+\,\color{red}{12H_2O}\,+\,}光エネルギー\\ \\
\,\hspace{7pt}\mathrm{\rightarrow \,C_6H_{12}O_6\,+\,\color{green}{6H_2O}\,+\,6O_2}\)
ここで注意して欲しいのは、
反応系の \(\mathrm{\color{red}{12H_2O}}\) は光化学系Ⅱにおいて水素イオンと電子をつくり出し、酸素を気孔を通じて排出するのに利用され、
生成系の \(\mathrm{\color{green}{6H_2O}}\) はカルビン・ベンソン回路でPGAがGAPに還元されるときに生成する水であるということです。
化学反応式としては反応系と生成系で差し引いて
\(\mathrm{6CO_2\,+\,6H_2O\,+\,}光エネルギー\\ \\
\,\hspace{7pt}\mathrm{\rightarrow \,C_6H_{12}O_6\,+\,6O_2}\)
と書いても良さそうなものですが、
違う水を示すので反応式ではあえて別物として書かれているのです。
C4、CAM植物の光合成
\(\mathrm{C_4}\)植物植物の中でもサトウキビやトウモロコシは、カルビン・ベンソン回路以外にも二酸化炭素を \(\mathrm{C_4}\) 化合物に一時的に蓄えることができます。
そのため \(\mathrm{C_4}\) 植物と呼ばれることがあるのですが、この \(\mathrm{C_4}\) 回路のおかげで効率よく二酸化炭素を固定することができます。
この \(\mathrm{C_4}\) 回路では葉肉細胞の葉緑体で、オキサロ酢酸と二酸化炭素が結合し、リンゴ酸となり固定され、維管束の周囲の維管束鞘(いかんそくしょう)細胞に送られてカルビン・ベンソン回路で有機物に合成されます。
CAM植物その他に、砂漠地帯で水分が蒸発しやすい昼間は気孔を開かず夜間に気孔を開いて二酸化炭素を吸収して蓄える仕組みをベンケイソウ型代謝を持つサボテンのような植物をCAM植物といいます。
CAM植物では二酸化炭素を夜間にリンゴ酸などに固定し液胞に蓄えておき、昼間になってからカルビン・ベンソン回路に送り出し有機物に合成する、という時間差同化を行います。
\(\mathrm{C_4}\) 植物は場所による分離で速度を落とすことなく光合成を効率的に行い、
CAM植物では時間的な分離(時間差)で水分を失わず光合成を効率的に行っているといえます。
これらの機能を持つようになったことは素晴らしいとしかいいようがないですね。
呼吸と光合成の共通点と違い
呼吸は異化、簡単にいえば分解で、
光合成は同化、つまり合成です。
逆の反応なので共通点はあるのかという疑問も出てきますが、
生体内では反応を起こすエネルギー(活性化エネルギー)をATPから得ますので共通点はあります。
呼吸の場であるミトコンドリアのクリステにある電子伝達系と、
⇒ 呼吸(解糖系・クエン酸回路・電子伝達系)で必要な酵素としくみ
光合成の場である葉緑体のチラコイドの電子伝達系の仕組みは非常によく似ています。
クリステとチラコイドの膜上に存在しているATP合成酵素はよく似ているし、水素イオンの流れを利用しATPを生成する仕組みもよく似ています。
ただ、呼吸では酸化的リン酸化といいますが、光合成では光リン酸化といいます。
まとめ
光合成は葉緑体で行われる。
光化学系の反応中心にある色素はクロロフィルである。
チラコイドで光エネルギーを使ってATPを合成する過程を光リン酸化という。
葉緑体で生成する酸素は「水」由来の酸素である。
葉緑体で二酸化炭素を固定する反応系をカルビン・ベンソン回路という。
呼吸と光合成の共通点は、
電子伝達系のしくみ
ATP合成のしくみ
ATP合成酵素の構造
がよく似ている点である。
以上、光合成の反応、仕組みをまとめて見ました。
光合成は1つの場所で行われる反応ではありません。
全体的な動きをある程度つかんでおくとイメージし易いです。
細胞小器官の中の一部での反応でもあるので非常に小さな場所でイメージしにくいのですが、
葉緑体の構造モデルで、物質の出入りだけでも覚えておくと良いですね。
光合成を行う細菌もいます。